

Volume 16, Issue 2 (March & April 2025)
BCN 2025, 16(2): 461-474 |
Back to browse issues page



Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Abroodi H, Joghataei M T, Shekari E, Nilipour R, Valinejad V, Almasi Doghaee M. Specific Brain Regions Involved in Decoding of the Anger Acoustic Parameters. BCN 2025; 16 (2) :461-474
URL: http://bcn.iums.ac.ir/article-1-2871-en.html
URL: http://bcn.iums.ac.ir/article-1-2871-en.html
Hossein Abroodi *1 

 , Mohammad Taghi Joghataei2 , Ehsan Shekari1 , Reza Nilipour3 , Vahid Valinejad4 , Mostafa Almasi Doghaee5
, Mohammad Taghi Joghataei2 , Ehsan Shekari1 , Reza Nilipour3 , Vahid Valinejad4 , Mostafa Almasi Doghaee5
, Mohammad Taghi Joghataei2 , Ehsan Shekari1 , Reza Nilipour3 , Vahid Valinejad4 , Mostafa Almasi Doghaee5
1- Department of Neuroscience, School of Advanced Technologies in Medicine, Iran University of Medical Sciences, Tehran, Iran.
2- Department of Innovation in Medical Education, Faculty of Medicine, Ottawa University, Ontario, Canada.
3- Department of Speech Therapy, University of Social Welfare and Rehabilitation Sciences, Tehran, Iran.
4- Department of Speech-Language Pathology, School of Rehabilitation, Tehran University of Medical Sciences, Tehran, Iran
5- Department of Neurology, Firoozgar Hospital, Iran University of Medical Sciences, Tehran, Iran.
2- Department of Innovation in Medical Education, Faculty of Medicine, Ottawa University, Ontario, Canada.
3- Department of Speech Therapy, University of Social Welfare and Rehabilitation Sciences, Tehran, Iran.
4- Department of Speech-Language Pathology, School of Rehabilitation, Tehran University of Medical Sciences, Tehran, Iran
5- Department of Neurology, Firoozgar Hospital, Iran University of Medical Sciences, Tehran, Iran.
Keywords: Acoustic parameters, Emotional prosody, Functional magnetic resonance imaging (fMRI), Intensity, Mean F0, Speech tempo
Full-Text [PDF 1412 kb]
| Abstract (HTML)
Full-Text:
1. Introduction
Speech prosody refers to fluctuations in pitch (fundamental frequency), variations in loudness (intensity), durational features (e.g. phone, syllable, word, and phrase length; pausing, rhythm, and speech tempo), and voice quality (Aziz-Zadeh et al., 2010; Mitchell & Ross, 2008; Rood et al., 2009; Sidtis & Van Lancker Sidtis, 2003). Prosody can serve various linguistic, pragmatic, and emotional functions (Lucarini et al., 2020). Affective states can influence these parameters and play an important role in emotion perception (Truong & Van Leeuwen, 2007). An incorrect interpretation of emotional prosody can cause failure in social interactions and an increased risk of social isolation (Durfee et al., 2021; Koch et al., 2018; Struchen et al., 2011). Finding the neural mechanism underlying the emotion-specific acoustic parameters can enrich our knowledge of prosodic impairments (dysprosodia).
Many neurological studies have found that the right cerebral cortex is responsible for emotional prosody processing (Alba-Ferrara et al., 2012; Behrens, 1989; Blonder et al., 1991; Blonder et al., 1995; Brådvik et al., 1991; Dara et al., 2014; Demenescu et al., 2015; Elizalde Acevedo et al., 2022; Geigenberger & Ziegler, 2001; Gibson et al., 2016; Grandjean, 2021; Grandjean et al., 2008; Heilman et al., 1984; Hoekert et al., 2010; Jacob et al., 2014; Kucharska-Pietura et al., 2003; Pell, 1999; Pell & Baum, 1997; Ross & Monnot, 2008, Ross & Monnot, 2011; Ryalls et al., 1987; Schmidt, 2003; Seydell-Greenwald et al., 2020; Shapiro & Danly, 1985; Sherratt, 2007; Tompkins & Flowers, 1985; Witteman et al., 2011; Wright et al., 2018; Wright et al., 2016). Although most lesions and neurological studies have suggested the right hemisphere (RH) dominance for emotional prosody, others have found no difference between the left hemisphere (LH) and RH lesion effects (House et al., 1987; Pell, 2006; Pell & Baum, 1997; Schlanger et al., 1976; Seydell-Greenwald et al., 2020; Twist et al., 1991).
Functional magnetic resonance imaging (fMRI) studies have showed widespread brain networks decode emotional prosody (Brück et al., 2011; Frühholz et al., 2015; Witteman et al., 2012). In detail, auditory temporal regions such as the primary/secondary auditory cortex (AC), superior temporal cortex (STC) (Alba-Ferrara et al., 2012; Beaucousin et al., 2007; Brück et al., 2011; Buchanan et al., 2000; Dietrich et al., 2008; Ethofer et al., 2006; Ethofer et al., 2012; Frühholz et al., 2012; Grandjean et al., 2005; Kotz et al., 2013; Kotz et al., 2003; Leitman et al., 2010; Mitchell et al., 2003; Mothes-Lasch et al., 2011; Sander et al., 2005; Schirmer & Kotz, 2006; Wiethoff et al., 2008; Witteman et al., 2012; Wittfoth et al., 2010), and other temporal areas such as supramarginal gyrus, right parahippocampal (BA 28) gyrus and subcallosal (BA 34) gyrus (Belyk & Brown, 2014), frontal regions such as the inferior frontal cortex (IFC) and orbital frontal cortex (OFC) (Brück et al., 2011; Ethofer et al., 2009; Frühholz et al., 2012; Kotz et al., 2013; Kotz et al., 2003; Leitman et al., 2010; Mitchell et al., 2003; Morris et al., 1999), insula (Mothes-Lasch et al., 2011), cerebellum (Belyk & Brown, 2014), and subcortical structures, such as thalamus, basal ganglia, and amygdala (Ethofer et al., 2009; Fecteau et al., 2007; Kotz et al., 2003; Leitman et al., 2010; Morris et al., 1999; Mothes-Lasch et al., 2011; Wiethoff et al., 2008) are involved in the perception of emotional prosody.
Furthermore, according to a hierarchical model that has been proposed for emotional prosody processing (Bach et al., 2008; Brück et al., 2011; Ethofer et al., 2006), voice-sensitive structures of the AC and mid-STC contribute to the extraction of acoustic parameters; the posterior part of the right STC contributes to the identification of emotional prosody, and eventually, the semantic comprehension of affective prosody is concerned with the bilateral inferior frontal gyrus (IFG) and OFC (Schirmer & Kotz, 2006; Witteman et al., 2012).
As discussed above, most studies have focused on emotional prosody processing, and there are few reports on how emotion-specific acoustic parameters are processed. We refer to studies on separately processing acoustic parameters, including pitch, intensity, and duration.
Pitch
The lesion studies addressed the role of the hemispheres in pitch processing. It has been demonstrated that RH is related to pitch processing in speech (Robin et al., 1990). Patients with RH lesions use duration cues rather than F0-variability to assess affective prosody. In other words, right temporoparietal lesions could disrupt the discrimination of tones (Robin et al., 1990).
Several structural and functional neuroimaging studies have shown that inferior frontal regions in RH (Gandour, 2000; Gandour et al., 1998), left superior temporal gyrus (STG), left Heschl’s gyrus (HG), and the right temporal pole (Wiethoff et al., 2008), pars triangularis of Broca’s area (Nan & Friederici, 2013), the right posterior STG and left STG subregions (Frühholz et al., 2015), HG, and adjacent cortical areas in STG (Warren et al., 2005) were the main structures for pitch processing. In addition, it has been suggested that the lateral HG functions as a general “pitch center” (Bendor & Wang, 2006), and the processing of pitch patterns, such as melodies, involves much more distributed processing in the superior temporal lobes and frontal lobes (Griffiths, 2003). However, another study demonstrated that parts of the planum temporal are more relevant for pitch processing than lateral HG (Hall & Plack, 2009). In addition, the anterior temporal cortex is more sensitive to female voices with high F0 than male voices with low F0 (Sokhi et al., 2005).
Intensity and duration
Previous reports found widespread activation clusters in the bilateral AC to stimuli that are deviant in intensity or duration (Langers et al., 2007; Mathiak et al., 2002; Schall et al., 2003), in the STG and middle temporal gyrus (MTG) of both hemispheres to intensity and duration, in the right IFG and superior frontal gyrus (SFG) to duration (Wiethoff et al., 2008), and in the right posterior STG and the left STG to intensity variations in emotional prosody (Frühholz et al., 2015; Warren et al., 2006).
In sum, different brain regions may be involved in processing emotion-specific acoustic parameters. Finding the neural representation of each emotion-specific acoustic parameter may improve our knowledge about the neural mechanism of emotional prosody processing. Difficulties in emotional prosody comprehension, including anger in neurological disorders such as Alzheimer, Parkinson, traumatic brain injury, or psychological disorders such as depression, autism, and alexithymia cause failure in social relationships and increase the risk of isolation (Durfee et al., 2021; Koch et al., 2018; Struchen et al., 2011). Recent studies have demonstrated improvement in some neurological, psychological, and motor disorders using non-invasive protocols such as tDCS (transcranial direct current stimulation) and rTMS (repetitive transcranial magnetic stimulation). Previous studies showed that the stimulation of a region such as the dorsal and lateral part of the prefrontal cortex leads to the strengthening of the functional connectivity of the brain networks (Clarke et al., 2020; Nitsche et al., 2012). Rare studies have been conducted in the treatment of emotional prosody disorders. Findings that the brain regions and functional connectivity are sensitive to changing the acoustic parameters of the emotional prosody of anger can help provide therapeutic solutions.
The emotional prosody used in the present study was anger. It was demonstrated in the previous reports (Sobin & Alpert, 1999) that lower mean F0, louder voice, and faster speech tempo constitute acoustic parameters of anger. However, it was revealed in one study (Banse & Scherer, 1996) that “hot” anger seemed to be characterized by an increase in mean F0, and a decrease in mean F0 was probably due to “cold” anger. Thus, it is expected that with increasing intensity and speech tempo and changing the mean F0, the anger becomes more intense, and the brain regions sensitive to these parameters show different activity. In this study, we tried to use a novel experimental fMRI design in which a range of varying anger stimuli with differences in only one acoustic parameter were presented to participants every time.
The goals of the present study were two-fold. In the first place, we aimed to identify brain regions sensitive to emotion-specific acoustic parameters. In the second place, we wanted to investigate the difference in brain activity related to emotion-specific acoustic parameter variations. We hypothesized that increasing anger intensity following lower frequency, louder (more intensity), and faster speech tempo leads to stronger activity in the brain regions sensitive to emotion-specific acoustic parameters. To ensure the relevance of the results with the difference in only one emotion-specific acoustic parameter, we changed one of the acoustic parameters each time while keeping the other two constants, and the stimuli were spoken only by one speech and language pathologist.
2. Materials and Methods
Study participants
Twenty healthy young male adults participated in the study (ages 18 to 35, Mean±SD age: 23.51±5.08 years). All participants were native Persian speakers, right-handed, and had normal or corrected-to-normal vision. No participant had a history of neurological or psychiatric problems, substance abuse, or impaired hearing. Furthermore, the Toronto alexithymia scale was used to identify individuals with trouble understanding emotions. All participants provided informed and written consent for participation in the fMRI study. MRI data had to be excluded from four participants because of incorrect responses. Thus, the results reported are based on an analysis of the remaining 16 participants.
Stimuli
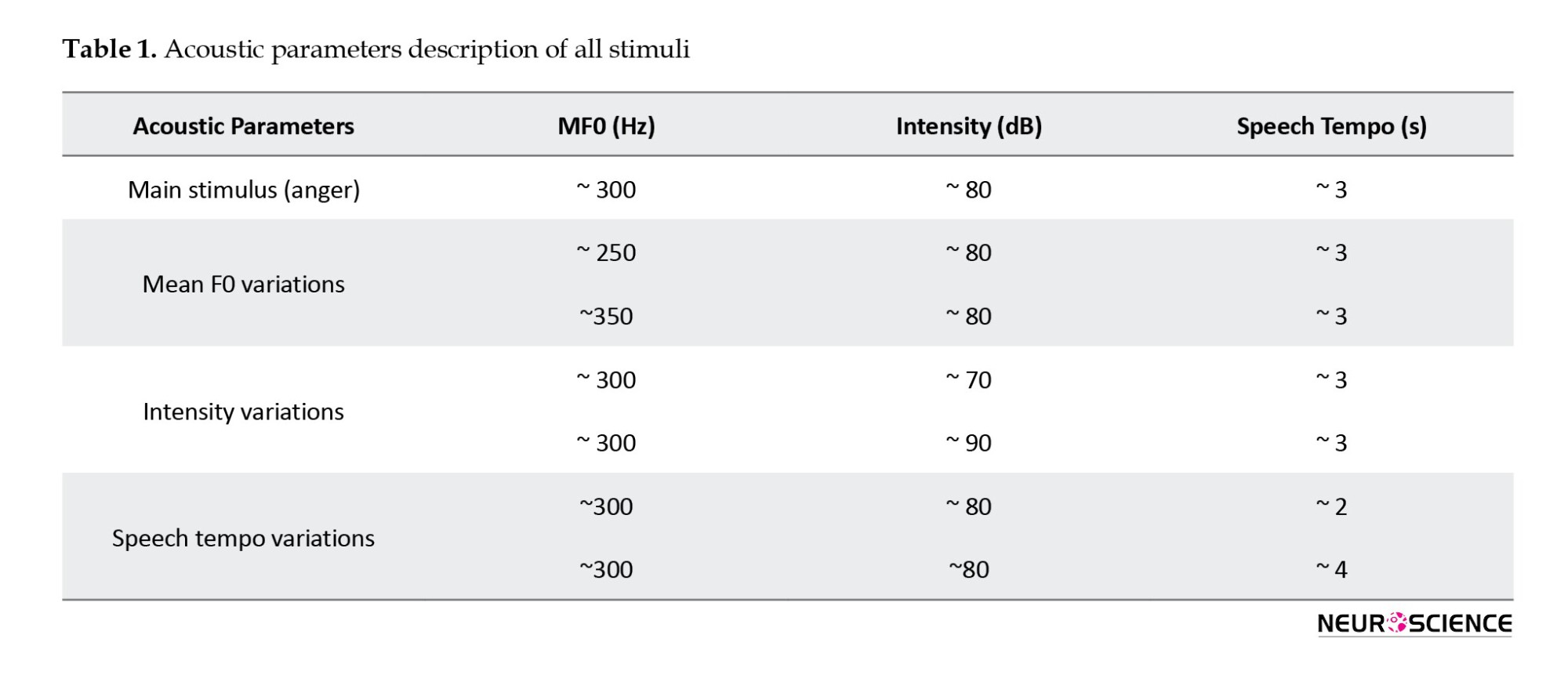
Three pseudo-words (“čârs,” “mâruk,” and “nirâpat”) were selected from a Persian language study (Kazemi & Saeednia, 2017) that was administered a Persian non-word repetition (NWR) test. A male speech and language pathologist spoke these pseudo-words in neutral or angry tones. After that, the three pseudo-words were used consecutively as one stimulus. Then, the acoustic parameters (mean F0, intensity, and speech tempo) of the anger stimulus were changed using Audacity software, version 2.1.0. Every time, one of the parameters was changed while keeping the other two constant. In detail, the mean F0 was changed between 200 and 400 Hz while keeping the other two parameters constant. Once again, keeping the mean F0 and tempo constant changed the mean intensity between 50 and 90 dB. Finally, the speech tempo was changed between 1 to 5 seconds without changing the mean F0 and intensity. After applying the changes to judge whether they were angry, they were piloted on a group of healthy adults (n=10) before the fMRI study. These healthy individuals were selected among the Iran University of Medical Sciences students. Finally, two changes were chosen in each acoustic parameter’s highest and lowest range of anger. The highest and lowest mean F0 detectable as anger prosody was about 350 and 250 Hz, respectively. The highest and lowest rate of change in speech tempo that could be recognized as anger prosody was about 4.2 and 2.1 seconds, respectively. The fMRI’s highest and lowest audible intensity were about 90 and 70 dBs, respectively. Therefore, 6 new changes (two changes in each parameter) were administrated (Table 1).
Experimental design
Auditory stimuli were presented binaurally during the fMRI scan using magnetic resonance imaging compatible headphones. The participants were lying in the scanner with their eyes open, staring at the screen. Auditory stimuli were preceded by a visual fixation cross (1×1°) for 2 s. The participants were asked to pay attention to the emotion of the auditory stimuli in the fMRI experiment to detect anger or neutral stimulus and distinguish the correct option by pressing the button (right index, left index) after displaying the options on the screen for 2 s. Auditory stimuli were presented during three blocks of prosody discrimination on the stimuli (angry or neutral; right index and left index) in two runs. Each anger/neutral discrimination block contained 12 conditions with 3 parameters (anger_neutral, stimulus_time, jitter), including 2 silent events with no auditory stimulation, 3 neutral events, and 7 anger events (Figure 1). Every block of prosody discrimination on the stimuli lasted 132 seconds.

Image acquisition
Structural and functional imaging data were obtained using a 3T PRISMA scanner in the National Brain Mapping Laboratory (NBML). A magnetization-prepared rapid acquisition gradient echo (MPRAGE) m3 sequence was employed to acquire high-resolution (1×1×1 mm3) T1-weighted structural images (TR [prepetition time]=1600 ms, TE [echo time]=3.47 ms, TI [inversion time]=800 ms). Functional images were obtained using a multislice echo planar imaging (EPI) sequence (36 axial slices, slice thickness: 3.0 mm, TR=2000 ms, TE=30 ms, field of view (FOV)=195 mm, flip angle=90°).
Image analysis
The fMRI data analysis used statistical parametric mapping (SPM; version 12; Welcome Department of Cognitive Neurology, London, UK). Preprocessing was performed using default settings in SPM 12. Functional images were realigned and coregistered to the anatomical image. A segmentation of the anatomical image revealed warping parameters that were used to normalize the functional images to the Montreal Neurological Institute (MNI) stereotactic template brain. Normalized images were spatially smoothed with a non-isotropic Gaussian kernel of full width at half-maximum 3×3×4 mm3.
A general linear model (Friston et al., 1994) was used for the first-level statistical analysis, in which separate regressors were defined for each trial using a stick function convolved with the hemodynamic response function. Events were time-locked to stimulus onset. Separate regressors were created for each experimental condition. Linear contrasts for the conditions for each participant were taken to a second-level random effects group analysis of variance.
Factorial subtraction analysis evaluated which brain regions respond more strongly to changes in emotion-specific acoustic parameters of anger prosody. To examine whether hemodynamic responses in the temporal lobe, especially STG, are subject to repetition suppression effects, parameter estimates of the most significantly activated voxel in this area were submitted to a two-factorial analysis of variance (ANOVA) with emotion-specific acoustic parameters variations and repetition (first fMRI session, second fMRI session) as within-subject factors. Activations are reported at a height threshold of P<0.001 uncorrected and an extent threshold of 0 voxels. Significance was examined at the cluster level with an extent threshold of P<0.05 (corresponding to a minimal cluster size of 50 voxels) corrected for multiple comparisons across the whole brain.
The instruction was displayed for 10 s before every block. A fixation cross was shown after a 5-7 s blank screen. After that, an audio stimulus was played to address anger/neutral discrimination. The duration of each stimulus was between about 2 and 4 seconds. The time required to respond was 2 seconds.
Results
The hemodynamic responses of the various acoustic parameters of anger prosody will be described in detail.
Mean F0
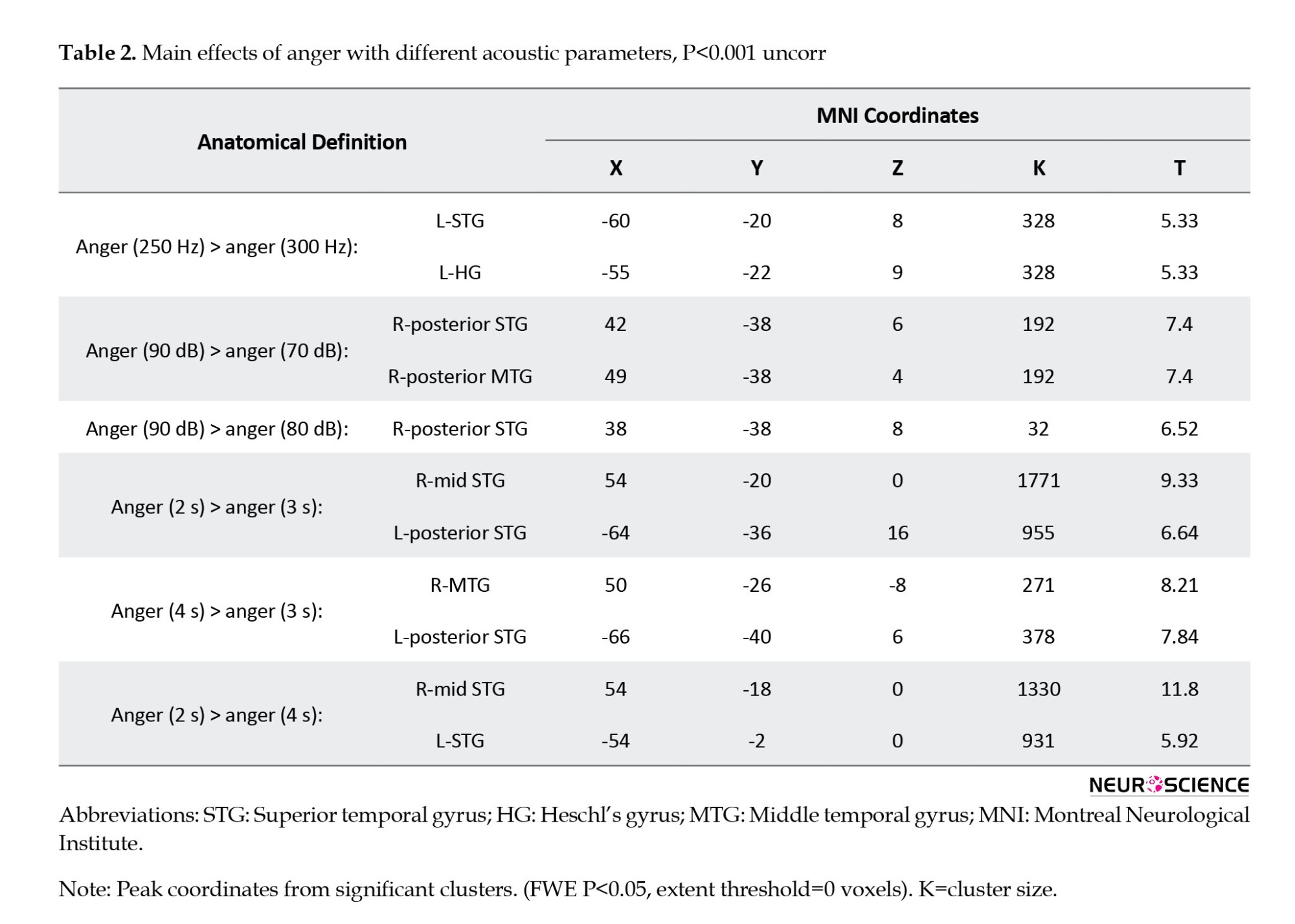
To examine brain activation in response to mean F0 variations in anger prosody, a 1-sample t-test was run for the contrasts: Anger (300 Hz) > anger (250 Hz) prosody, anger (250 Hz) > anger (300 Hz) prosody, anger (350 Hz) > anger (300 Hz) prosody, and anger (300 Hz) > anger (350 Hz) prosody. Comparison of these contrasts with each other showed increased activity in the left STG (MNI coordinates: x=-60, y=-20, z=8; T=5.33; cluster size=17 voxels) and in HG (MNI coordinates: x=-55, y=-22, z=9; T=5.33; cluster size=17 voxels) in response to anger (250 Hz) prosody compared with anger (300 Hz) at P<0.001 (uncorrected) (Figure 2; Table 2). No significant clusters were found in other contrast comparisons.

Mean intensity
When we compared brain activity among intensity variations (70, 80, and 90 dBs) while keeping the other two acoustic parameters constant, the STG and MTG in the RH showed greater activation to anger prosody with more intensity. This increased activation was greater for anger (90 dB) compared with anger (70 dB) [90 dB > 70 dB] than for anger (90 dB) compared with anger (80 dB) [90 dB > 80 dB] (Figure 2). In detail, when we compared brain activation between anger (90 dB) and anger (70dB) (90 dB >70 dB), our analysis revealed stronger activity in the right posterior STG (MNI coordinates: x=42, y=-38, z=6; T=7.4; cluster size=22 voxels) and a significant cluster in the right posterior MTG (MNI coordinates: x=49, y=-38, z=4; T=7.4; cluster size=22 voxels) at the threshold of P<0.001 uncorrected. In addition, when we compared brain activation between anger (90 dB) and anger (80 dB) (90 dB >80 dB), we found increased activity in the right posterior STG (MNI coordinates: x=38, y=-38, z=8; T=6.52; cluster size=21 voxels). Furthermore, the difference in activity extent in anger 90 dB compared with anger 70 dB is greater than anger 90 dB compared with anger 80 dB (Figure 2).
Duration (speech tempo)
Comparison of hemodynamic responses to anger (2 s) compared with anger (3 s) (anger, 2 s > anger, 3 s) revealed two significant clusters in the temporal lobe of both hemispheres, which were located within the mid-STG in the RH (MNI coordinates: x=54; y=-20; z=0; BA:22; T=9.3; cluster size=27 voxels), and the posterior STG in the LH (MNI coordinates: x=-64; y=-36; z=16; BA:22; T=6.6; cluster size=27 voxels) at the threshold of P<0.001 (uncorrected) with a minimum cluster extent of k=0 voxels. This activation was stronger in the RH (Figure 2). On the other hand, when we compared brain activity between anger (4 s) and anger (3 s) (anger, 4s > anger, 3s), we found increased activity in the right MTG (MNI coordinates: x=50; y=-26; z=-8; T=8.2; cluster size=21 voxels), and in the posterior STG in LH (MNI coordinates: x=-66; y=-40; z=6; T=7.8; cluster size=21 voxels) at the level of P<0.001 (uncorrected) with a minimum cluster extent of k=0 voxels. This activation was stronger in LH (Figure 2). Eventually, the comparison between anger (2 s) and anger (4 s) (anger, 2s > anger, 4s) revealed stronger activity in the right mid-STG (MNI coordinates: x=54; y=-18; z=0; T=11.8; cluster size=27 voxels) and in the left STG (MNI coordinates: x=-54; y=-2; z=0; T=5.9; cluster size=27 voxels) at the level of P<0.001 (uncorrected) (Figure 2).
Discussion
The present study investigated the brain activity underlying the decoding of emotion-specific acoustic parameters in anger prosody. We wanted to identify brain regions sensitive to changing acoustic parameters. According to our assumption, changing any emotion-specific acoustic parameters can result in different hemodynamic responses within the STG and MTG of both hemispheres. Our findings show stronger activation in both hemispheres’ STG, HG, and MTG when the acoustic parameters of anger are changed. Our results confirm previous reports on regions showing increased responsiveness to acoustic parameters (Beaucousin et al., 2007; Sander et al., 2005; Wiethoff et al., 2008). In previous studies, it has been suggested that the decoding of acoustic parameters occurs in the temporal lobe, especially in STG subregions, and there is no study on how brain activity differs in the temporal lobe following changes in any of these acoustic parameters. Therefore, we tried to change any acoustic parameters separately, keep the other two parameters constant, and examine how the brain activity differs.
When we changed the mean F0 from 300 Hz to 250 Hz and kept the intensity and duration constant, our analysis revealed stronger activity in HG and adjacent areas in STG in LH. The activity in the left STG in our findings is consistent with the results of previous studies (Frühholz et al., 2015; Wiethoff et al., 2008).
We also showed the role of HG in pitch processing, which agrees with the studies suggesting HG as a “pitch center” (Bendor & Wang, 2006). Previous studies (Banse & Scherer, 1996) demonstrated that one of the characteristics of “hot” anger was increased in mean F0. Studies in which this feature was not found may have been measuring “cold” anger. Our finding about mean F0 variations was probably due to the use of “cold” anger. In fact, according to the previous study (Sobin & Alpert, 1999), low fundamental frequency is one of the characteristics of anger. Anger is perceived as more intense, with a further decrease in the fundamental frequency. Therefore, we expected increased activity in the regions related to fundamental frequency decoding by reducing the mean F0. Thus, according to the use of “cold” anger in the present study, the increase in the mean F0 did not increase in the intensity of anger, and as expected, the brain regions related to mean F0 decoding should not show increased activity.
The present study observed no activity in the anterior temporal cortex. In fact, due to the use of male voices for the stimuli, we did not expect that the anterior temporal cortex would be active because, according to previous studies (Sokhi et al., 2005), the anterior temporal cortex is more sensitive to female voices with high F0 than to male voices with low F0. Thus, the inactivity of the anterior temporal cortex was predictable.
Our findings on the speech tempo revealed stronger activity in different parts of the STG and MTG of both hemispheres when we only changed the speech tempo. These findings align with previous studies’ results (Mathiak et al., 2002; Schall et al., 2003; Wiethoff et al., 2008), demonstrating activation clusters in the bilateral auditory cortex, including STG, to stimuli that are deviant in duration. Because the temporal lobe, especially STG, is mentioned in all studies on duration processing, we can address the importance of these regions in decoding duration. The slight difference in our results and those of previous studies probably goes back to the difference in the method and the type of acoustic parameter investigation. We examined the speech tempo as a durational feature, whereas others used duration-deviant tone in noise or stimuli duration in emotional prosody.
The results of our study about the effect of mean intensity variety on brain activity showed that increasing the mean intensity of anger prosody leads to increased activity in the posterior part of STG and MTG in the RH. The activity of the right posterior part of STG in our study agrees with previous findings (Frühholz et al., 2015; Warren et al., 2006). Inspection of responses in the right posterior MTG revealed a reasonably linear relationship within the investigated intensity range. These findings converge with previous findings demonstrating a linear relationship between BOLD (blood oxygenation level dependent) response and sound intensity for a range of intensity (Langers et al., 2007; Wiethoff et al., 2008). The type of stimulus used in our study and previous studies led to different results. In our study, only a kind of emotional prosody (anger) was used while keeping other parameters (mean F0, duration, voice gender) constant to investigate the effect of mean intensity variations on brain region activity. Other studies used frequency-modulated tones (Langers et al., 2007; or types of emotional prosody (Wiethoff et al., 2008). The latter study found widespread activity in different brain regions, including STG and MTG of both hemispheres, due to different types of emotional prosody without considering stimuli’s fundamental frequency and duration. At the same time, our results were limited to the posterior part of STG and MTG in the RH, probably due to using only anger prosody and keeping other parameters constant.
The STG and MTG subregions significantly correlated with changes in mean intensity, fundamental frequency, and duration (speech tempo). This sensitivity to various acoustic parameters is probably due to the role of STG subregions in the detailed analysis of voice information (Warren et al., 2005; Warren et al., 2006).
Conclusion
As discussed in the introduction, AC and mid-STC (Bach et al., 2008; Brück et al., 2011; Ethofer et al., 2006) have a role in the extraction of acoustic parameters and different parts of STG and MTG of both hemispheres in our findings in decoding the anger acoustic parameters. So, we can conclude that more intense anger following the lower fundamental frequency, increased intensity, and faster speech tempo leads to increased activity in the specific brain regions related to decoding the anger acoustic parameters. Eventually, different information is sent to IFG and OFC (Schirmer & Kotz, 2006; Witteman et al., 2012) for more intense comprehension of anger.
Study limitation and strength
The current study aimed to investigate the automatic processing of anger acoustic parameters. Thus, the experimental design did not include any behavioral tasks. Therefore, our study limitation is the lack of behavioral control, indicating that subjects comprehended the auditory information in the presence of scanner noise. However, none of the subjects reported difficulties in understanding the presented stimuli, and all subjects reported that all stimuli were spoken in an emotional tone of voice after scanning. On the other hand, to avoid the influence of various factors on the results of the study, including the use of various emotional prosody, different lengths of stimuli, presenting stimuli with the voices of different individuals, and the long duration of the task, we decided to use only one emotional prosody (anger) stimulus, change its acoustic parameters, and be spoken by only one male speech and language pathologist. Considering the different acoustic characteristics of emotional prosody and the distinct role of each acoustic parameter in the semantic comprehension of emotional prosody, we suggest that other emotional prosody should be investigated to improve our knowledge about decoding emotion-specific acoustic parameters.
Ethical Considerations
Compliance with ethical guidelines
This study was approved by the Ethics Committee of the Iran University of Medical Sciences, Tehran, Iran (Code: IR.IUMS.REC.1399.1284).
Funding
This study was supported by a grant from Iran University of Medical Sciences, Tehran, Iran.
Authors' contributions
Conceptualization, methodology, and software: Hossein Abroodi, and Reza Nilipour; Formal analysis: Hossein Abroodi, Ehsan Shekari and Vahid Valinejad; Investigation and resources: Hossein Abroodi, Mostafa Almasi Doghaee; Supervision: Mohammad Taghi Joghataei; Project administration: Hossein Abroodi; Funding acquisition: Mostafa Almasi Doghaee; Writing the original draft: Hossein Abroodi; Review and editing: All Authors.
Conflict of interest
The authors declared no conflict of interest.
Acknowledgments
The authors want to thank the National Brain Mapping Laboratory for their help in the implementation stages of the study.
References
Speech prosody refers to fluctuations in pitch (fundamental frequency), variations in loudness (intensity), durational features (e.g. phone, syllable, word, and phrase length; pausing, rhythm, and speech tempo), and voice quality (Aziz-Zadeh et al., 2010; Mitchell & Ross, 2008; Rood et al., 2009; Sidtis & Van Lancker Sidtis, 2003). Prosody can serve various linguistic, pragmatic, and emotional functions (Lucarini et al., 2020). Affective states can influence these parameters and play an important role in emotion perception (Truong & Van Leeuwen, 2007). An incorrect interpretation of emotional prosody can cause failure in social interactions and an increased risk of social isolation (Durfee et al., 2021; Koch et al., 2018; Struchen et al., 2011). Finding the neural mechanism underlying the emotion-specific acoustic parameters can enrich our knowledge of prosodic impairments (dysprosodia).
Many neurological studies have found that the right cerebral cortex is responsible for emotional prosody processing (Alba-Ferrara et al., 2012; Behrens, 1989; Blonder et al., 1991; Blonder et al., 1995; Brådvik et al., 1991; Dara et al., 2014; Demenescu et al., 2015; Elizalde Acevedo et al., 2022; Geigenberger & Ziegler, 2001; Gibson et al., 2016; Grandjean, 2021; Grandjean et al., 2008; Heilman et al., 1984; Hoekert et al., 2010; Jacob et al., 2014; Kucharska-Pietura et al., 2003; Pell, 1999; Pell & Baum, 1997; Ross & Monnot, 2008, Ross & Monnot, 2011; Ryalls et al., 1987; Schmidt, 2003; Seydell-Greenwald et al., 2020; Shapiro & Danly, 1985; Sherratt, 2007; Tompkins & Flowers, 1985; Witteman et al., 2011; Wright et al., 2018; Wright et al., 2016). Although most lesions and neurological studies have suggested the right hemisphere (RH) dominance for emotional prosody, others have found no difference between the left hemisphere (LH) and RH lesion effects (House et al., 1987; Pell, 2006; Pell & Baum, 1997; Schlanger et al., 1976; Seydell-Greenwald et al., 2020; Twist et al., 1991).
Functional magnetic resonance imaging (fMRI) studies have showed widespread brain networks decode emotional prosody (Brück et al., 2011; Frühholz et al., 2015; Witteman et al., 2012). In detail, auditory temporal regions such as the primary/secondary auditory cortex (AC), superior temporal cortex (STC) (Alba-Ferrara et al., 2012; Beaucousin et al., 2007; Brück et al., 2011; Buchanan et al., 2000; Dietrich et al., 2008; Ethofer et al., 2006; Ethofer et al., 2012; Frühholz et al., 2012; Grandjean et al., 2005; Kotz et al., 2013; Kotz et al., 2003; Leitman et al., 2010; Mitchell et al., 2003; Mothes-Lasch et al., 2011; Sander et al., 2005; Schirmer & Kotz, 2006; Wiethoff et al., 2008; Witteman et al., 2012; Wittfoth et al., 2010), and other temporal areas such as supramarginal gyrus, right parahippocampal (BA 28) gyrus and subcallosal (BA 34) gyrus (Belyk & Brown, 2014), frontal regions such as the inferior frontal cortex (IFC) and orbital frontal cortex (OFC) (Brück et al., 2011; Ethofer et al., 2009; Frühholz et al., 2012; Kotz et al., 2013; Kotz et al., 2003; Leitman et al., 2010; Mitchell et al., 2003; Morris et al., 1999), insula (Mothes-Lasch et al., 2011), cerebellum (Belyk & Brown, 2014), and subcortical structures, such as thalamus, basal ganglia, and amygdala (Ethofer et al., 2009; Fecteau et al., 2007; Kotz et al., 2003; Leitman et al., 2010; Morris et al., 1999; Mothes-Lasch et al., 2011; Wiethoff et al., 2008) are involved in the perception of emotional prosody.
Furthermore, according to a hierarchical model that has been proposed for emotional prosody processing (Bach et al., 2008; Brück et al., 2011; Ethofer et al., 2006), voice-sensitive structures of the AC and mid-STC contribute to the extraction of acoustic parameters; the posterior part of the right STC contributes to the identification of emotional prosody, and eventually, the semantic comprehension of affective prosody is concerned with the bilateral inferior frontal gyrus (IFG) and OFC (Schirmer & Kotz, 2006; Witteman et al., 2012).
As discussed above, most studies have focused on emotional prosody processing, and there are few reports on how emotion-specific acoustic parameters are processed. We refer to studies on separately processing acoustic parameters, including pitch, intensity, and duration.
Pitch
The lesion studies addressed the role of the hemispheres in pitch processing. It has been demonstrated that RH is related to pitch processing in speech (Robin et al., 1990). Patients with RH lesions use duration cues rather than F0-variability to assess affective prosody. In other words, right temporoparietal lesions could disrupt the discrimination of tones (Robin et al., 1990).
Several structural and functional neuroimaging studies have shown that inferior frontal regions in RH (Gandour, 2000; Gandour et al., 1998), left superior temporal gyrus (STG), left Heschl’s gyrus (HG), and the right temporal pole (Wiethoff et al., 2008), pars triangularis of Broca’s area (Nan & Friederici, 2013), the right posterior STG and left STG subregions (Frühholz et al., 2015), HG, and adjacent cortical areas in STG (Warren et al., 2005) were the main structures for pitch processing. In addition, it has been suggested that the lateral HG functions as a general “pitch center” (Bendor & Wang, 2006), and the processing of pitch patterns, such as melodies, involves much more distributed processing in the superior temporal lobes and frontal lobes (Griffiths, 2003). However, another study demonstrated that parts of the planum temporal are more relevant for pitch processing than lateral HG (Hall & Plack, 2009). In addition, the anterior temporal cortex is more sensitive to female voices with high F0 than male voices with low F0 (Sokhi et al., 2005).
Intensity and duration
Previous reports found widespread activation clusters in the bilateral AC to stimuli that are deviant in intensity or duration (Langers et al., 2007; Mathiak et al., 2002; Schall et al., 2003), in the STG and middle temporal gyrus (MTG) of both hemispheres to intensity and duration, in the right IFG and superior frontal gyrus (SFG) to duration (Wiethoff et al., 2008), and in the right posterior STG and the left STG to intensity variations in emotional prosody (Frühholz et al., 2015; Warren et al., 2006).
In sum, different brain regions may be involved in processing emotion-specific acoustic parameters. Finding the neural representation of each emotion-specific acoustic parameter may improve our knowledge about the neural mechanism of emotional prosody processing. Difficulties in emotional prosody comprehension, including anger in neurological disorders such as Alzheimer, Parkinson, traumatic brain injury, or psychological disorders such as depression, autism, and alexithymia cause failure in social relationships and increase the risk of isolation (Durfee et al., 2021; Koch et al., 2018; Struchen et al., 2011). Recent studies have demonstrated improvement in some neurological, psychological, and motor disorders using non-invasive protocols such as tDCS (transcranial direct current stimulation) and rTMS (repetitive transcranial magnetic stimulation). Previous studies showed that the stimulation of a region such as the dorsal and lateral part of the prefrontal cortex leads to the strengthening of the functional connectivity of the brain networks (Clarke et al., 2020; Nitsche et al., 2012). Rare studies have been conducted in the treatment of emotional prosody disorders. Findings that the brain regions and functional connectivity are sensitive to changing the acoustic parameters of the emotional prosody of anger can help provide therapeutic solutions.
The emotional prosody used in the present study was anger. It was demonstrated in the previous reports (Sobin & Alpert, 1999) that lower mean F0, louder voice, and faster speech tempo constitute acoustic parameters of anger. However, it was revealed in one study (Banse & Scherer, 1996) that “hot” anger seemed to be characterized by an increase in mean F0, and a decrease in mean F0 was probably due to “cold” anger. Thus, it is expected that with increasing intensity and speech tempo and changing the mean F0, the anger becomes more intense, and the brain regions sensitive to these parameters show different activity. In this study, we tried to use a novel experimental fMRI design in which a range of varying anger stimuli with differences in only one acoustic parameter were presented to participants every time.
The goals of the present study were two-fold. In the first place, we aimed to identify brain regions sensitive to emotion-specific acoustic parameters. In the second place, we wanted to investigate the difference in brain activity related to emotion-specific acoustic parameter variations. We hypothesized that increasing anger intensity following lower frequency, louder (more intensity), and faster speech tempo leads to stronger activity in the brain regions sensitive to emotion-specific acoustic parameters. To ensure the relevance of the results with the difference in only one emotion-specific acoustic parameter, we changed one of the acoustic parameters each time while keeping the other two constants, and the stimuli were spoken only by one speech and language pathologist.
2. Materials and Methods
Study participants
Twenty healthy young male adults participated in the study (ages 18 to 35, Mean±SD age: 23.51±5.08 years). All participants were native Persian speakers, right-handed, and had normal or corrected-to-normal vision. No participant had a history of neurological or psychiatric problems, substance abuse, or impaired hearing. Furthermore, the Toronto alexithymia scale was used to identify individuals with trouble understanding emotions. All participants provided informed and written consent for participation in the fMRI study. MRI data had to be excluded from four participants because of incorrect responses. Thus, the results reported are based on an analysis of the remaining 16 participants.
Stimuli
Three pseudo-words (“čârs,” “mâruk,” and “nirâpat”) were selected from a Persian language study (Kazemi & Saeednia, 2017) that was administered a Persian non-word repetition (NWR) test. A male speech and language pathologist spoke these pseudo-words in neutral or angry tones. After that, the three pseudo-words were used consecutively as one stimulus. Then, the acoustic parameters (mean F0, intensity, and speech tempo) of the anger stimulus were changed using Audacity software, version 2.1.0. Every time, one of the parameters was changed while keeping the other two constant. In detail, the mean F0 was changed between 200 and 400 Hz while keeping the other two parameters constant. Once again, keeping the mean F0 and tempo constant changed the mean intensity between 50 and 90 dB. Finally, the speech tempo was changed between 1 to 5 seconds without changing the mean F0 and intensity. After applying the changes to judge whether they were angry, they were piloted on a group of healthy adults (n=10) before the fMRI study. These healthy individuals were selected among the Iran University of Medical Sciences students. Finally, two changes were chosen in each acoustic parameter’s highest and lowest range of anger. The highest and lowest mean F0 detectable as anger prosody was about 350 and 250 Hz, respectively. The highest and lowest rate of change in speech tempo that could be recognized as anger prosody was about 4.2 and 2.1 seconds, respectively. The fMRI’s highest and lowest audible intensity were about 90 and 70 dBs, respectively. Therefore, 6 new changes (two changes in each parameter) were administrated (Table 1).
Experimental design
Auditory stimuli were presented binaurally during the fMRI scan using magnetic resonance imaging compatible headphones. The participants were lying in the scanner with their eyes open, staring at the screen. Auditory stimuli were preceded by a visual fixation cross (1×1°) for 2 s. The participants were asked to pay attention to the emotion of the auditory stimuli in the fMRI experiment to detect anger or neutral stimulus and distinguish the correct option by pressing the button (right index, left index) after displaying the options on the screen for 2 s. Auditory stimuli were presented during three blocks of prosody discrimination on the stimuli (angry or neutral; right index and left index) in two runs. Each anger/neutral discrimination block contained 12 conditions with 3 parameters (anger_neutral, stimulus_time, jitter), including 2 silent events with no auditory stimulation, 3 neutral events, and 7 anger events (Figure 1). Every block of prosody discrimination on the stimuli lasted 132 seconds.
Image acquisition
Structural and functional imaging data were obtained using a 3T PRISMA scanner in the National Brain Mapping Laboratory (NBML). A magnetization-prepared rapid acquisition gradient echo (MPRAGE) m3 sequence was employed to acquire high-resolution (1×1×1 mm3) T1-weighted structural images (TR [prepetition time]=1600 ms, TE [echo time]=3.47 ms, TI [inversion time]=800 ms). Functional images were obtained using a multislice echo planar imaging (EPI) sequence (36 axial slices, slice thickness: 3.0 mm, TR=2000 ms, TE=30 ms, field of view (FOV)=195 mm, flip angle=90°).
Image analysis
The fMRI data analysis used statistical parametric mapping (SPM; version 12; Welcome Department of Cognitive Neurology, London, UK). Preprocessing was performed using default settings in SPM 12. Functional images were realigned and coregistered to the anatomical image. A segmentation of the anatomical image revealed warping parameters that were used to normalize the functional images to the Montreal Neurological Institute (MNI) stereotactic template brain. Normalized images were spatially smoothed with a non-isotropic Gaussian kernel of full width at half-maximum 3×3×4 mm3.
A general linear model (Friston et al., 1994) was used for the first-level statistical analysis, in which separate regressors were defined for each trial using a stick function convolved with the hemodynamic response function. Events were time-locked to stimulus onset. Separate regressors were created for each experimental condition. Linear contrasts for the conditions for each participant were taken to a second-level random effects group analysis of variance.
Factorial subtraction analysis evaluated which brain regions respond more strongly to changes in emotion-specific acoustic parameters of anger prosody. To examine whether hemodynamic responses in the temporal lobe, especially STG, are subject to repetition suppression effects, parameter estimates of the most significantly activated voxel in this area were submitted to a two-factorial analysis of variance (ANOVA) with emotion-specific acoustic parameters variations and repetition (first fMRI session, second fMRI session) as within-subject factors. Activations are reported at a height threshold of P<0.001 uncorrected and an extent threshold of 0 voxels. Significance was examined at the cluster level with an extent threshold of P<0.05 (corresponding to a minimal cluster size of 50 voxels) corrected for multiple comparisons across the whole brain.
The instruction was displayed for 10 s before every block. A fixation cross was shown after a 5-7 s blank screen. After that, an audio stimulus was played to address anger/neutral discrimination. The duration of each stimulus was between about 2 and 4 seconds. The time required to respond was 2 seconds.
Results
The hemodynamic responses of the various acoustic parameters of anger prosody will be described in detail.
Mean F0
To examine brain activation in response to mean F0 variations in anger prosody, a 1-sample t-test was run for the contrasts: Anger (300 Hz) > anger (250 Hz) prosody, anger (250 Hz) > anger (300 Hz) prosody, anger (350 Hz) > anger (300 Hz) prosody, and anger (300 Hz) > anger (350 Hz) prosody. Comparison of these contrasts with each other showed increased activity in the left STG (MNI coordinates: x=-60, y=-20, z=8; T=5.33; cluster size=17 voxels) and in HG (MNI coordinates: x=-55, y=-22, z=9; T=5.33; cluster size=17 voxels) in response to anger (250 Hz) prosody compared with anger (300 Hz) at P<0.001 (uncorrected) (Figure 2; Table 2). No significant clusters were found in other contrast comparisons.
Mean intensity
When we compared brain activity among intensity variations (70, 80, and 90 dBs) while keeping the other two acoustic parameters constant, the STG and MTG in the RH showed greater activation to anger prosody with more intensity. This increased activation was greater for anger (90 dB) compared with anger (70 dB) [90 dB > 70 dB] than for anger (90 dB) compared with anger (80 dB) [90 dB > 80 dB] (Figure 2). In detail, when we compared brain activation between anger (90 dB) and anger (70dB) (90 dB >70 dB), our analysis revealed stronger activity in the right posterior STG (MNI coordinates: x=42, y=-38, z=6; T=7.4; cluster size=22 voxels) and a significant cluster in the right posterior MTG (MNI coordinates: x=49, y=-38, z=4; T=7.4; cluster size=22 voxels) at the threshold of P<0.001 uncorrected. In addition, when we compared brain activation between anger (90 dB) and anger (80 dB) (90 dB >80 dB), we found increased activity in the right posterior STG (MNI coordinates: x=38, y=-38, z=8; T=6.52; cluster size=21 voxels). Furthermore, the difference in activity extent in anger 90 dB compared with anger 70 dB is greater than anger 90 dB compared with anger 80 dB (Figure 2).
Duration (speech tempo)
Comparison of hemodynamic responses to anger (2 s) compared with anger (3 s) (anger, 2 s > anger, 3 s) revealed two significant clusters in the temporal lobe of both hemispheres, which were located within the mid-STG in the RH (MNI coordinates: x=54; y=-20; z=0; BA:22; T=9.3; cluster size=27 voxels), and the posterior STG in the LH (MNI coordinates: x=-64; y=-36; z=16; BA:22; T=6.6; cluster size=27 voxels) at the threshold of P<0.001 (uncorrected) with a minimum cluster extent of k=0 voxels. This activation was stronger in the RH (Figure 2). On the other hand, when we compared brain activity between anger (4 s) and anger (3 s) (anger, 4s > anger, 3s), we found increased activity in the right MTG (MNI coordinates: x=50; y=-26; z=-8; T=8.2; cluster size=21 voxels), and in the posterior STG in LH (MNI coordinates: x=-66; y=-40; z=6; T=7.8; cluster size=21 voxels) at the level of P<0.001 (uncorrected) with a minimum cluster extent of k=0 voxels. This activation was stronger in LH (Figure 2). Eventually, the comparison between anger (2 s) and anger (4 s) (anger, 2s > anger, 4s) revealed stronger activity in the right mid-STG (MNI coordinates: x=54; y=-18; z=0; T=11.8; cluster size=27 voxels) and in the left STG (MNI coordinates: x=-54; y=-2; z=0; T=5.9; cluster size=27 voxels) at the level of P<0.001 (uncorrected) (Figure 2).
Discussion
The present study investigated the brain activity underlying the decoding of emotion-specific acoustic parameters in anger prosody. We wanted to identify brain regions sensitive to changing acoustic parameters. According to our assumption, changing any emotion-specific acoustic parameters can result in different hemodynamic responses within the STG and MTG of both hemispheres. Our findings show stronger activation in both hemispheres’ STG, HG, and MTG when the acoustic parameters of anger are changed. Our results confirm previous reports on regions showing increased responsiveness to acoustic parameters (Beaucousin et al., 2007; Sander et al., 2005; Wiethoff et al., 2008). In previous studies, it has been suggested that the decoding of acoustic parameters occurs in the temporal lobe, especially in STG subregions, and there is no study on how brain activity differs in the temporal lobe following changes in any of these acoustic parameters. Therefore, we tried to change any acoustic parameters separately, keep the other two parameters constant, and examine how the brain activity differs.
When we changed the mean F0 from 300 Hz to 250 Hz and kept the intensity and duration constant, our analysis revealed stronger activity in HG and adjacent areas in STG in LH. The activity in the left STG in our findings is consistent with the results of previous studies (Frühholz et al., 2015; Wiethoff et al., 2008).
We also showed the role of HG in pitch processing, which agrees with the studies suggesting HG as a “pitch center” (Bendor & Wang, 2006). Previous studies (Banse & Scherer, 1996) demonstrated that one of the characteristics of “hot” anger was increased in mean F0. Studies in which this feature was not found may have been measuring “cold” anger. Our finding about mean F0 variations was probably due to the use of “cold” anger. In fact, according to the previous study (Sobin & Alpert, 1999), low fundamental frequency is one of the characteristics of anger. Anger is perceived as more intense, with a further decrease in the fundamental frequency. Therefore, we expected increased activity in the regions related to fundamental frequency decoding by reducing the mean F0. Thus, according to the use of “cold” anger in the present study, the increase in the mean F0 did not increase in the intensity of anger, and as expected, the brain regions related to mean F0 decoding should not show increased activity.
The present study observed no activity in the anterior temporal cortex. In fact, due to the use of male voices for the stimuli, we did not expect that the anterior temporal cortex would be active because, according to previous studies (Sokhi et al., 2005), the anterior temporal cortex is more sensitive to female voices with high F0 than to male voices with low F0. Thus, the inactivity of the anterior temporal cortex was predictable.
Our findings on the speech tempo revealed stronger activity in different parts of the STG and MTG of both hemispheres when we only changed the speech tempo. These findings align with previous studies’ results (Mathiak et al., 2002; Schall et al., 2003; Wiethoff et al., 2008), demonstrating activation clusters in the bilateral auditory cortex, including STG, to stimuli that are deviant in duration. Because the temporal lobe, especially STG, is mentioned in all studies on duration processing, we can address the importance of these regions in decoding duration. The slight difference in our results and those of previous studies probably goes back to the difference in the method and the type of acoustic parameter investigation. We examined the speech tempo as a durational feature, whereas others used duration-deviant tone in noise or stimuli duration in emotional prosody.
The results of our study about the effect of mean intensity variety on brain activity showed that increasing the mean intensity of anger prosody leads to increased activity in the posterior part of STG and MTG in the RH. The activity of the right posterior part of STG in our study agrees with previous findings (Frühholz et al., 2015; Warren et al., 2006). Inspection of responses in the right posterior MTG revealed a reasonably linear relationship within the investigated intensity range. These findings converge with previous findings demonstrating a linear relationship between BOLD (blood oxygenation level dependent) response and sound intensity for a range of intensity (Langers et al., 2007; Wiethoff et al., 2008). The type of stimulus used in our study and previous studies led to different results. In our study, only a kind of emotional prosody (anger) was used while keeping other parameters (mean F0, duration, voice gender) constant to investigate the effect of mean intensity variations on brain region activity. Other studies used frequency-modulated tones (Langers et al., 2007; or types of emotional prosody (Wiethoff et al., 2008). The latter study found widespread activity in different brain regions, including STG and MTG of both hemispheres, due to different types of emotional prosody without considering stimuli’s fundamental frequency and duration. At the same time, our results were limited to the posterior part of STG and MTG in the RH, probably due to using only anger prosody and keeping other parameters constant.
The STG and MTG subregions significantly correlated with changes in mean intensity, fundamental frequency, and duration (speech tempo). This sensitivity to various acoustic parameters is probably due to the role of STG subregions in the detailed analysis of voice information (Warren et al., 2005; Warren et al., 2006).
Conclusion
As discussed in the introduction, AC and mid-STC (Bach et al., 2008; Brück et al., 2011; Ethofer et al., 2006) have a role in the extraction of acoustic parameters and different parts of STG and MTG of both hemispheres in our findings in decoding the anger acoustic parameters. So, we can conclude that more intense anger following the lower fundamental frequency, increased intensity, and faster speech tempo leads to increased activity in the specific brain regions related to decoding the anger acoustic parameters. Eventually, different information is sent to IFG and OFC (Schirmer & Kotz, 2006; Witteman et al., 2012) for more intense comprehension of anger.
Study limitation and strength
The current study aimed to investigate the automatic processing of anger acoustic parameters. Thus, the experimental design did not include any behavioral tasks. Therefore, our study limitation is the lack of behavioral control, indicating that subjects comprehended the auditory information in the presence of scanner noise. However, none of the subjects reported difficulties in understanding the presented stimuli, and all subjects reported that all stimuli were spoken in an emotional tone of voice after scanning. On the other hand, to avoid the influence of various factors on the results of the study, including the use of various emotional prosody, different lengths of stimuli, presenting stimuli with the voices of different individuals, and the long duration of the task, we decided to use only one emotional prosody (anger) stimulus, change its acoustic parameters, and be spoken by only one male speech and language pathologist. Considering the different acoustic characteristics of emotional prosody and the distinct role of each acoustic parameter in the semantic comprehension of emotional prosody, we suggest that other emotional prosody should be investigated to improve our knowledge about decoding emotion-specific acoustic parameters.
Ethical Considerations
Compliance with ethical guidelines
This study was approved by the Ethics Committee of the Iran University of Medical Sciences, Tehran, Iran (Code: IR.IUMS.REC.1399.1284).
Funding
This study was supported by a grant from Iran University of Medical Sciences, Tehran, Iran.
Authors' contributions
Conceptualization, methodology, and software: Hossein Abroodi, and Reza Nilipour; Formal analysis: Hossein Abroodi, Ehsan Shekari and Vahid Valinejad; Investigation and resources: Hossein Abroodi, Mostafa Almasi Doghaee; Supervision: Mohammad Taghi Joghataei; Project administration: Hossein Abroodi; Funding acquisition: Mostafa Almasi Doghaee; Writing the original draft: Hossein Abroodi; Review and editing: All Authors.
Conflict of interest
The authors declared no conflict of interest.
Acknowledgments
The authors want to thank the National Brain Mapping Laboratory for their help in the implementation stages of the study.
References
Alba-Ferrara, L., Ellison, A., & Mitchell, R. L. C. (2012). Decoding emotional prosody: resolving differences in functional neuroanatomy from fMRI and lesion studies using TMS. Brain Stimulation, 5(3), 347–353. [DOI:10.1016/j.brs.2011.06.004] [PMID]
Aziz-Zadeh, L., Sheng, T., & Gheytanchi, A. (2010). Common premotor regions for the perception and production of prosody and correlations with empathy and prosodic ability. Plos One, 5(1), e8759. [DOI:10.1371/journal.pone.0008759] [PMID]
Bach, D. R., Grandjean, D., Sander, D., Herdener, M., Strik, W. K., & Seifritz, E. (2008). The effect of appraisal level on processing of emotional prosody in meaningless speech. Neuroimage, 42(2), 919-927. [DOI:10.1016/j.neuroimage.2008.05.034] [PMID]
Banse, R., & Scherer, K. R. (1996). Acoustic profiles in vocal emotion expression. Journal of Personality and Social Psychology, 70(3), 614–636. [DOI:10.1037//0022-3514.70.3.614] [PMID]
Beaucousin, V., Lacheret, A., Turbelin, M. R., Morel, M., Mazoyer, B., & Tzourio-Mazoyer, N. (2007). FMRI study of emotional speech comprehension. Cerebral Cortex (New York, N.Y. : 1991), 17(2), 339–352. [DOI:10.1093/cercor/bhj151] [PMID]
Behrens, S. J. (1989). Characterizing sentence intonation in a right hemisphere-damaged population. Brain and Language, 37(2), 181–200. [DOI:10.1016/0093-934x(89)90014-x] [PMID]
Belyk, M., & Brown, S. (2014). Perception of affective and linguistic prosody: An ALE meta-analysis of neuroimaging studies. Social Cognitive and Affective Neuroscience, 9(9), 1395–1403. [DOI:10.1093/scan/nst124] [PMID]
Bendor, D., & Wang, X. (2006). Cortical representations of pitch in monkeys and humans. Current Opinion in Neurobiology, 16(4), 391–399. [DOI:10.1016/j.conb.2006.07.001] [PMID]
Blonder, L. X., Bowers, D., & Heilman, K. M. (1991). The role of the right hemisphere in emotional communication. Brain, 114 ( Pt 3), 1115-1127. [DOI:10.1093/brain/114.3.1115] [PMID]
Blonder, L. X., Pickering, J. E., Heath, R. L., Smith, C. D., & Butler, S. M. (1995). Prosodic characteristics of speech pre- and post-right hemisphere stroke. Brain and Language, 51(2), 318–335. [DOI:10.1006/brln.1995.1063] [PMID]
Brådvik, B., Dravins, C., Holtås, S., Rosén, I., Ryding, E., & Ingvar, D. H. (1991). Disturbances of speech prosody following right hemisphere infarcts. Acta Neurologica Scandinavica, 84(2), 114–126. [DOI:10.1111/j.1600-0404.1991.tb04919.x] [PMID]
Brück, C., Kreifelts, B., & Wildgruber, D. (2011). Emotional voices in context: A neurobiological model of multimodal affective information processing. Physics of Life Reviews, 8(4), 383–403. [DOI:10.1016/j.plrev.2011.10.002] [PMID]
Buchanan, T. W., Lutz, K., Mirzazade, S., Specht, K., Shah, N. J., & Zilles, K., et al. (2000). Recognition of emotional prosody and verbal components of spoken language: An fMRI study. Brain Research. Cognitive Brain Research, 9(3), 227–238. [DOI:10.1016/s0926-6410(99)00060-9] [PMID]
Clarke, P. J. F., Van Bockstaele, B., Marinovic, W., Howell, J. A., Boyes, M. E., & Notebaert, L. (2020). The effects of left DLPFC tDCS on emotion regulation, biased attention, and emotional reactivity to negative content. Cognitive, Affective & Behavioral Neuroscience, 20(6), 1323–1335. [DOI:10.3758/s13415-020-00840-2] [PMID]
Dara, C., Bang, J., Gottesman, R. F., & Hillis, A. E. (2014). Right hemisphere dysfunction is better predicted by emotional prosody impairments as compared to neglect. Journal of Neurology & Translational Neuroscience, 2(1), 1037. [PMID]
Demenescu, L. R., Kato, Y., & Mathiak, K. (2015). Neural processing of emotional prosody across the adult lifespan. BioMed Research International, 2015, 590216. [DOI:10.1155/2015/590216] [PMID]
Dietrich, S., Hertrich, I., Alter, K., Ischebeck, A., & Ackermann, H. (2008). Understanding the emotional expression of verbal interjections: a functional MRI study. Neuroreport, 19(18), 1751–1755. [DOI:10.1097/WNR.0b013e3283193e9e] [PMID]
Durfee, A. Z., Sheppard, S. M., Blake, M. L., & Hillis, A. E. (2021). Lesion loci of impaired affective prosody: A systematic review of evidence from stroke. Brain and Cognition, 152, 105759. [DOI:10.1016/j.bandc.2021.105759] [PMID]
Elizalde Acevedo, B., Olano, M. A., Bendersky, M., Kochen, S., Agüero Vera, V., & Chambeaud, N., et al. (2022). Brain mapping of emotional prosody in patients with drug-resistant temporal epilepsy: An indicator of plasticity. Cortex; A Journal Devoted to The Study of the Nervous System and Behavior, 153, 97–109. [DOI:10.1016/j.cortex.2022.04.014] [PMID]
Ethofer, T., Anders, S., Erb, M., Herbert, C., Wiethoff, S., & Kissler, J., et al. (2006). Cerebral pathways in processing of affective prosody: A dynamic causal modeling study. NeuroImage, 30(2), 580–587. [DOI:10.1016/j.neuroimage.2005.09.059] [PMID]
Ethofer, T., Bretscher, J., Gschwind, M., Kreifelts, B., Wildgruber, D., & Vuilleumier, P. (2012). Emotional voice areas: Anatomic location, functional properties, and structural connections revealed by combined fMRI/DTI. Cerebral Cortex (New York, N.Y. : 1991), 22(1), 191–200. [DOI:10.1093/cercor/bhr113] [PMID]
Ethofer, T., Van De Ville, D., Scherer, K., & Vuilleumier, P. (2009). Decoding of emotional information in voice-sensitive cortices. Current Biology : CB, 19(12), 1028–1033. [DOI:10.1016/j.cub.2009.04.054] [PMID]
Fecteau, S., Belin, P., Joanette, Y., & Armony, J. L. (2007). Amygdala responses to nonlinguistic emotional vocalizations. Neuroimage, 36(2), 480-487. [DOI:10.1016/j.neuroimage.2007.02.043] [PMID]
Friston, K. J., Holmes, A. P., Worsley, K. J., Poline, J. P., Frith, C. D., & Frackowiak, R. S. (1994). Statistical parametric maps in functional imaging: A general linear approach. Human Brain Mapping, 2(4), 189-210. [DOI:10.1002/hbm.460020402]
Frühholz, S., Ceravolo, L., & Grandjean, D. (2012). Specific brain networks during explicit and implicit decoding of emotional prosody. Cerebral Cortex (New York, N.Y. : 1991), 22(5), 1107–1117. [DOI:10.1093/cercor/bhr184] [PMID]
Frühholz, S., Gschwind, M., & Grandjean, D. (2015). Bilateral dorsal and ventral fiber pathways for the processing of affective prosody identified by probabilistic fiber tracking. Neuroimage, 109, 27-34. [DOI:10.1016/j.neuroimage.2015.01.016] [PMID]
Gandour, J. (2000). Frontiers of brain mapping of speech prosody. Brain and Language, 71(1), 75–77. [DOI:10.1006/brln.1999.2217] [PMID]
Gandour, J., Wong, D., & Hutchins, G. (1998). Pitch processing in the human brain is influenced by language experience. Neuroreport, 9(9), 2115-2119. [DOI:10.1097/00001756-199806220-00038] [PMID]
Geigenberger, A., & Ziegler, W. (2001). The processing of prosodic patterns in patients with unilateral brain lesions. Clinical Linguistics & Phonetics, 15(1-2), 85–89. [DOI:10.3109/02699200109167636] [PMID]
Gibson, L., Atchley, R. A., Voyer, D., Diener, U. S., & Gregersen, S. (2016). Detection of sarcastic speech: The role of the right hemisphere in ambiguity resolution. Laterality, 21(4-6), 549-567. [DOI:10.1080/1357650x.2015.1105246] [PMID]
Grandjean, D. (2020). Brain networks of emotional prosody processing. Emotion Review, 13(1), 34-43. [DOI:10.1177/1754073919898522]
Grandjean, D., Sander, D., Lucas, N., Scherer, K. R., & Vuilleumier, P. (2008). Effects of emotional prosody on auditory extinction for voices in patients with spatial neglect. Neuropsychologia, 46(2), 487-496. [DOI:10.1016/j.neuropsychologia.2007.08.025] [PMID]
Grandjean, D., Sander, D., Pourtois, G., Schwartz, S., Seghier, M. L., & Scherer, K. R., et al. (2005). The voices of wrath: Brain responses to angry prosody in meaningless speech. Nature Neuroscience, 8(2), 145–146. [DOI:10.1038/nn1392] [PMID]
Griffiths, T. D. (2003). Functional imaging of pitch analysis. Annals of The New York Academy of Sciences, 999, 40–49. [DOI:10.1196/annals.1284.004] [PMID]
Hall, D. A., & Plack, C. J. (2009). Pitch processing sites in the human auditory brain. Cerebral Cortex (New York, N.Y. : 1991), 19(3), 576–585. [DOI:10.1093/cercor/bhn108] [PMID]
Heilman, K. M., Bowers, D., Speedie, L., & Coslett, H. B. (1984). Comprehension of affective and nonaffective prosody. Neurology, 34(7), 917-921. [DOI:10.1212/wnl.34.7.917] [PMID]
Hoekert, M., Vingerhoets, G., & Aleman, A. (2010). Results of a pilot study on the involvement of bilateral inferior frontal gyri in emotional prosody perception: An rTMS study. BMC Neuroscience, 11, 93. [DOI:10.1186/1471-2202-11-93] [PMID]
House, A., Rowe, D., & Standen, P. J. (1987). Affective prosody in the reading voice of stroke patients. Journal of Neurology, Neurosurgery, and Psychiatry, 50(7), 910–912. [DOI:10.1136/jnnp.50.7.910] [PMID]
Jacob, H., Brück, C., Plewnia, C., & Wildgruber, D. (2014). Cerebral processing of prosodic emotional signals: Evaluation of a network model using rTMS. Plos One, 9(8), e105509. [DOI:10.1371/journal.pone.0105509] [PMID]
Kazemi, Y., & Saeednia, S. (2017). The clinical examination of non-word repetition tasks in identifying Persian-speaking children with primary language impairment. International Journal of Pediatric Otorhinolaryngology, 93, 7–12. [DOI:10.1016/j.ijporl.2016.11.028] [PMID]
Koch, K., Stegmaier, S., Schwarz, L., Erb, M., Reinl, M., & Scheffler, K., et al. (2018). Neural correlates of processing emotional prosody in unipolar depression. Human Brain Mapping, 39(8), 3419–3427. [DOI:10.1002/hbm.24185] [PMID]
Kotz, S. A., Kalberlah, C., Bahlmann, J., Friederici, A. D., & Haynes, J. D. (2013). Predicting vocal emotion expressions from the human brain. Human Brain Mapping, 34(8), 1971–1981. [DOI:10.1002/hbm.22041] [PMID]
Kotz, S. A., Meyer, M., Alter, K., Besson, M., von Cramon, D. Y., & Friederici, A. D. (2003). On the lateralization of emotional prosody: An event-related functional MR investigation. Brain and Language, 86(3), 366–376. [DOI:10.1016/s0093-934x(02)00532-1] [PMID]
Kucharska-Pietura, K., Phillips, M. L., Gernand, W., & David, A. S. (2003). Perception of emotions from faces and voices following unilateral brain damage. Neuropsychologia, 41(8), 1082–1090. [DOI:10.1016/s0028-3932(02)00294-4] [PMID]
Langers, D. R., van Dijk, P., Schoenmaker, E. S., & Backes, W. H. (2007). fMRI activation in relation to sound intensity and loudness. Neuroimage, 35(2), 709-718. [DOI:10.1016/j.neuroimage.2006.12.013] [PMID]
Leitman, D. I., Wolf, D. H., Ragland, J. D., Laukka, P., Loughead, J., & Valdez, J. N., et al. (2010). "It's Not What You Say, But How You Say it": A Reciprocal Temporo-frontal Network for Affective Prosody. Frontiers in Human Neuroscience, 4, 19. [DOI:10.3389/fnhum.2010.00019] [PMID]
Lucarini, V., Grice, M., Cangemi, F., Zimmermann, J. T., Marchesi, C., & Vogeley, K., et al. (2020). Speech Prosody as a Bridge Between Psychopathology and Linguistics: The Case of the Schizophrenia Spectrum. Frontiers in Psychiatry, 11, 531863. [DOI:10.3389/fpsyt.2020.531863] [PMID]
Mathiak, K., Rapp, A., Kircher, T. T., Grodd, W., Hertrich, I., & Weiskopf, N., et al. (2002). Mismatch responses to randomized gradient switching noise as reflected by fMRI and whole-head magnetoencephalography. Human Brain Mapping, 16(3), 190–195. [DOI:10.1002/hbm.10041] [PMID]
Mitchell, R. L., Elliott, R., Barry, M., Cruttenden, A., & Woodruff, P. W. (2003). The neural response to emotional prosody, as revealed by functional magnetic resonance imaging. Neuropsychologia, 41(10), 1410–1421. [DOI:10.1016/s0028-3932(03)00017-4] [PMID]
Mitchell, R. L., & Ross, E. D. (2008). fMRI evidence for the effect of verbal complexity on lateralisation of the neural response associated with decoding prosodic emotion. Neuropsychologia, 46(12), 2880-2887. [DOI:10.1016/j.neuropsychologia.2008.05.024] [PMID]
Morris, J. S., Scott, S. K., & Dolan, R. J. (1999). Saying it with feeling: neural responses to emotional vocalizations. Neuropsychologia, 37(10), 1155-1163. [DOI:10.1016/s0028-3932(99)00015-9] [PMID]
Mothes-Lasch, M., Mentzel, H. J., Miltner, W. H., & Straube, T. (2011). Visual attention modulates brain activation to angry voices. The Journal of Neuroscience : The Official Journal of The Society for Neuroscience, 31(26), 9594–9598. [DOI:10.1523/jneurosci.6665-10.2011] [PMID]
Nan, Y., & Friederici, A. D. (2013). Differential roles of right temporal cortex and Broca’s area in pitch processing: Evidence from music and Mandarin. Human Brain Mapping, 34(9), 2045–2054. [DOI:10.1002/hbm.22046] [PMID]
Nitsche, M. A., Koschack, J., Pohlers, H., Hullemann, S., Paulus, W., & Happe, S. (2012). Effects of frontal transcranial direct current stimulation on emotional state and processing in healthy humans. Frontiers in Psychiatry, 3, 58. [DOI:10.3389/fpsyt.2012.00058] [PMID]
Pell, M. D. (1999). Fundamental frequency encoding of linguistic and emotional prosody by right hemisphere-damaged speakers. Brain and Language, 69(2), 161–192. [DOI:10.1006/brln.1999.2065] [PMID]
Pell, M. D. (2006). Cerebral mechanisms for understanding emotional prosody in speech. Brain and Language, 96(2), 221–234.[DOI:10.1016/j.bandl.2005.04.007] [PMID]
Pell, M. D., & Baum, S. R. (1997). Unilateral brain damage, prosodic comprehension deficits, and the acoustic cues to prosody. Brain and Language, 57(2), 195–214. [DOI:10.1006/brln.1997.1736] [PMID]
Robin, D. A., Tranel, D., & Damasio, H. (1990). Auditory perception of temporal and spectral events in patients with focal left and right cerebral lesions. Brain and Language, 39(4), 539–555.[DOI:10.1016/0093-934x(90)90161-9] [PMID]
Rood, L., Roelofs, J., Bögels, S. M., Nolen-Hoeksema, S., & Schouten, E. (2009). The influence of emotion-focused rumination and distraction on depressive symptoms in non-clinical youth: A meta-analytic review. Clinical Psychology Review, 29(7), 607–616. [DOI:10.1016/j.cpr.2009.07.001] [PMID]
Ross, E. D., & Monnot, M. (2008). Neurology of affective prosody and its functional-anatomic organization in right hemisphere. Brain and Language, 104(1), 51–74. [DOI:10.1016/j.bandl.2007.04.007] [PMID]
Ross, E. D., & Monnot, M. (2011). Affective prosody: what do comprehension errors tell us about hemispheric lateralization of emotions, sex and aging effects, and the role of cognitive appraisal. Neuropsychologia, 49(5), 866-877. [DOI:10.1016/j.neuropsychologia.2010.12.024] [PMID]
Ryalls, J., Joanette, Y., & Feldman, L. (1987). An acoustic comparison of normal and right-hemisphere-damaged speech prosody. Cortex, 23(4), 685-694. [DOI:10.1016/s0010-9452(87)80059-x] [PMID]
Sander, D., Grandjean, D., Pourtois, G., Schwartz, S., Seghier, M. L., & Scherer, K. R., et al. (2005). Emotion and attention interactions in social cognition: Brain regions involved in processing anger prosody. Neuroimage, 28(4), 848-858. [DOI:10.1016/j.neuroimage.2005.06.023] [PMID]
Schall, U., Johnston, P., Todd, J., Ward, P. B., & Michie, P. T. (2003). Functional neuroanatomy of auditory mismatch processing: An event-related fMRI study of duration-deviant oddballs. NeuroImage, 20(2), 729–736. [DOI:10.1016/s1053-8119(03)00398-7] [PMID]
Schirmer, A., & Kotz, S. A. (2006). Beyond the right hemisphere: Brain mechanisms mediating vocal emotional processing. Trends in Cognitive Sciences, 10(1), 24–30. [DOI:10.1016/j.tics.2005.11.009] [PMID]
Schlanger, B. B., Schlanger, P., & Gerstman, L. J. (1976). The perception of emotionally toned sentences by right hemisphere-damaged and aphasic subjects. Brain and Language, 3(3), 396–403.[DOI:10.1016/0093-934x(76)90035-3] [PMID]
Schmidt, J. M. (2003). Emotional prosody production in brain-damaged populations: Human judgments and acoustical analysis [PhD dissertation]. New York: City University of New York. [Link]
Seydell-Greenwald, A., Chambers, C. E., Ferrara, K., & Newport, E. L. (2020). What you say versus how you say it: Comparing sentence comprehension and emotional prosody processing using fMRI. Neuroimage, 209, 116509. [DOI:10.1016/j.neuroimage.2019.116509] [PMID]
Shapiro, B. E., & Danly, M. (1985). The role of the right hemisphere in the control of speech prosody in propositional and affective contexts. Brain and Language, 25(1), 19–36. [DOI:10.1016/0093-934x(85)90118-x] [PMID]
Sherratt, S. (2007). Right brain damage and the verbal expression of emotion: A preliminary investigation. Aphasiology, 21(3-4), 320-339. [DOI:10.1080/02687030600911401]
Sidtis, J. J., & Van Lancker Sidtis, D. (2003). A neurobehavioral approach to dysprosody. Seminars in Speech and Language, 24(2), 93–105. [DOI:10.1055/s-2003-38901] [PMID]
Sobin, C., & Alpert, M. (1999). Emotion in speech: The acoustic attributes of fear, anger, sadness, and joy. Journal of Psycholinguistic Research, 28(4), 347–365. [DOI:10.1023/a:1023237014909] [PMID]
Sokhi, D. S., Hunter, M. D., Wilkinson, I. D., & Woodruff, P. W. (2005). Male and female voices activate distinct regions in the male brain. Neuroimage, 27(3), 572-578. [DOI:10.1016/j.neuroimage.2005.04.023] [PMID]
Struchen, M. A., Pappadis, M. R., Sander, A. M., Burrows, C. S., & Myszka, K. A. (2011). Examining the contribution of social communication abilities and affective/behavioral functioning to social integration outcomes for adults with traumatic brain injury. The Journal of Head Trauma Rehabilitation, 26(1), 30–42. [DOI:10.1097/HTR.0b013e3182048f7c] [PMID]
Tompkins, C. A., & Flowers, C. R. (1985). Perception of emotional intonation by brain-damaged adults: The influence of task processing levels. Journal of Speech and Hearing Research, 28(4), 527–538. [DOI:10.1044/jshr.2804.527] [PMID]
Truong, K., & Van Leeuwen, D. (2007). Automatic discrimination between laughter and speech. Speech Communication, 49(2), 144-158. [DOI:10.1016/j.specom.2007.01.001]
Twist, D. J., Squires, N. K., Spielholz, N. I., & Silverglide, R. (1991). Event-related potentials in disorders of prosodic and semantic linguistic processing 1. Cognitive and Behavioral Neurology, 4(4), 281-304. [Link]
Warren, J. D., Jennings, A., & Griffiths, T. D. (2005). Analysis of the spectral envelope of sounds by the human brain. Neuroimage, 24(4), 1052-1057. [DOI:10.1016/j.neuroimage.2004.10.031] [PMID]
Warren, J. D., Scott, S. K., Price, C. J., & Griffiths, T. D. (2006). Human brain mechanisms for the early analysis of voices. NeuroImage, 31(3), 1389–1397. [DOI:10.1016/j.neuroimage.2006.01.034] [PMID]
Wiethoff, S., Wildgruber, D., Kreifelts, B., Becker, H., Herbert, C., Grodd, W., & Ethofer, T. (2008). Cerebral processing of emotional prosody--influence of acoustic parameters and arousal. NeuroImage, 39(2), 885–893. [DOI:10.1016/j.neuroimage.2007.09.028] [PMID]
Witteman, J., Van Heuven, V. J. P., & Schiller, N. O. (2012). Hearing feelings: A quantitative meta-analysis on the neuroimaging literature of emotional prosody perception. Neuropsychologia, 50(12), 2752–2763. [DOI:10.1016/j.neuropsychologia.2012.07.026] [PMID]
Witteman, J., van Ijzendoorn, M. H., van de Velde, D., van Heuven, V. J., & Schiller, N. O. (2011). The nature of hemispheric specialization for linguistic and emotional prosodic perception: a meta-analysis of the lesion literature. Neuropsychologia, 49(13), 3722–3738. [DOI:10.1016/j.neuropsychologia.2011.09.028] [PMID]
Wittfoth, M., Schröder, C., Schardt, D. M., Dengler, R., Heinze, H. J., & Kotz, S. A. (2010). On emotional conflict: Interference resolution of happy and angry prosody reveals valence-specific effects. Cerebral Cortex (New York, N.Y. : 1991), 20(2), 383–392. [DOI:10.1093/cercor/bhp106] [PMID]
Wright, A., Saxena, S., Sheppard, S. M., & Hillis, A. E. (2018). Selective impairments in components of affective prosody in neurologically impaired individuals. Brain and Cognition, 124, 29–36. [DOI:10.1016/j.bandc.2018.04.001] [PMID]
Wright, A. E., Davis, C., Gomez, Y., Posner, J., Rorden, C., & Hillis, A. E., et al. (2016). Acute ischemic lesions associated with impairments in expression and recognition of affective prosody. Perspectives of the ASHA Special Interest Groups, 1(2), 82–95. [DOI:10.1044/persp1.SIG2.82] [PMID]
Type of Study: Original |
Subject:
Cellular and molecular Neuroscience
Received: 2024/01/22 | Accepted: 2024/06/17 | Published: 2025/03/1
Received: 2024/01/22 | Accepted: 2024/06/17 | Published: 2025/03/1
Send email to the article author
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |
![]()
Copyright © The Author(s);
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC-By-NC), which permits use, distribution, and reproduction in any medium, provided the original work is properly cited and is not used for commercial purposes.
Contact Information
